
An Alternative Analysis on Nilsson-Ehle’s Hybridization Experiment in Wheat —Theory of Dual Multiple
Vol.09No.08(2018), Article ID:86993,10 pages
10.4236/am.2018.98068
―Theory of Dual Multiple Factors and Three Normal Distributions on Quantitative Inheritance (Continuation)
Tingzhen Zhang1*, Xiaoming Jia1, Zhao Xu2
1College of Forestry, Northwest A & F University, Yangling, China
2College of Science, Northwest A & F University, Yangling, China

Copyright © 2018 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: July 26, 2018; Accepted: August 27, 2018; Published: August 30, 2018
ABSTRACT
In this paper, an analysis on Nilsson-Ehle’s hybridization experiment in wheat was done by means of self-fertilization method along with the pericarp inheritance. It showed that because seeds of wheat were coated with 2n tissues of mother’s body, the grain colors were determined by mother’s genotype. The color of the F1 grains in this experiment was old red. The phenotypes of the F2 grains were uniformly medium red and did not segregate. Grains in the F3 generation segregated in the ratio 6 red (varying intensities of red) to 1 white with a definite probability respectively. This is the consequence of additive effect of 3 gene pairs and belongs to discrete distribution rather than continuous distribution. Therefore, the multiple-gene hypothesis based on this experiment cannot solve the continuous variations in inheritance of quantitative characters.
Keywords:
Wheat Hybridization, 3 Gene Pairs, Additive Effect, Grain Color, Pericarp Inheritance, Delayed Inheritance, Discrete Distribution

1. Introduction
In early 1900s Nilsson-Ehle conducted hybridization experiments in red seed variety and white seed variety in wheat. On the basis of this study, the famous multiple-gene hypothesis was founded and used to explain the continuity of quantitative characters by William Bateson, G. Udny Yule and other geneticists [1]. This hypothesis has become a classic theory of the inheritance of quantitative character [2]and has been popular so far over the world. In this study, an alternative analysis was done for this experiment and it pointed out the misjudgments of genetics community in the world and drawbacks of this hypothesis.
2. Data Sources and Analysis Methods
In 1900 Nilsson-Ehle found a wheat variety with old red seed coat in Northern Sweden and crossed it with a white seed variety later on. He got 78 F2 plants out of a total of 64 plants of the theoretical expectation. 8 segregating the F3 red plants and white plants in the ratio 3:1, 15 segregating the F3 red plants and white plants in the ratio 15:1, 5 segregating the F3 red plants and white plants in the ratio 63:1, the F3 of 50 plants were all red plants, the F3 of 0 plants were all white plants [3].
Among them the ratio 63:1 should be identical with the expected ratio of plants producing red grains and white grains in the F2 generation. This is the symbol of inheritance of 3 gene pairs.
Because seeds in wheat are coated with 2n tissues of mother’s body-pericarp [4][5], the grain colors rely on the pericarp inheritance and are determined by mother’s genotypes [6]. According to inheritance of 3 gene pairs, the above data would be tentatively analyzed by means of self-fertilization method along with the pericarp inheritance.
3. Genetic Analysis
3.1. The F2 Genotypes and Seed Phenotypes in Inheritance of 3 Gene Pairs in Crops
The hereditary manners in wheat and other crops are the same, but segregation timelines of their seed colors are different.
3.1.1. The F2 Genotypes and Their Categories, and Seed Phenotypes in the Inheritance of 3 Gene Pairs in Garden Pea
When red seed variety with 2 gene pairs was crossed with white seed variety in wheat, the variation in grain color was explained with the help of additive effect [7]. In additive effect, two kinds of dominant genes exist simultaneously and alone, characteristics controlled by them are similar [8]. Different reds in wheat grains are similar characteristics. If additive effect was be expanded to inheritance of 3 gene pairs, for example, the inheritance of 3 gene pairs in garden pea would be studied based on this effect. Regarding the results of Nilsson-Ehle’s experiment, according to the additive effect, the F2 genotypes from the inheritance of 3 gene pairs in garden pea [8]can be classified into the following 5 categories: 1) homozygotes with 3 pairs of recessive genes, 2) heterozygotes with 1 pair of heterozygous genes, 3) heterozygotes with 2 pairs of heterozygous genes, 4) heterozygotes with 3 pairs of heterozygous genes, 5) individuals containing 1 - 3 pairs of homozygous dominant genes, by adding one dominant gene at a time (Table 1).
Phenotypes of the F2 seed colors in garden pea: Dividing generation in crops begins with seeds. The seeds produced by maternal parents used in crossing and

Table 1. Categories of the F2 genotype from inheritance of 3 gene pairs in garden pea.
the plants grown from these seeds are the F1. The seeds produced by the F1 plants have been the starting point of the F2 [6][8]. The inheritance of 3 gene pairs in garden pea is known. The above analysis is only used to describe the classification of genotypes considering the additive effect from 3 gene pairs. In fact, the seed color in garden pea is independent inherited without interaction of genes. And the round yellow and wrinkled green of seeds in garden pea [9]are the traits of 2n tissue of embryo-cotyledon [8][10]. Phenotypes of the F2 seed colors are determined by the F2 genotypes and in the second hybrid generation the seed colors segregated 3 yellow and 1 green [9].
3.1.2. The F2 Genotypes, Their Categories and Grain Phenotypes in Crossing with Red Seed Wheat of 3 Gene Pairs
In 1909, Nilsson-Ehle used an old red seed variety as mother parent and crossed it with white seed variety [1][3][8]to produce the F1 old red grains, genotype of which is r1R1r2R2r3R3. These grains were sown, and the F1 plants grown from them produced medium red grains (F2) by self-fertilization [6](Figure 1).
As additive effect in garden pea mentioned above, the F2 genotypes in wheat are classified, so that the classification is consistent with the genotype category in garden pea. Because alleles at 3 loci in wheat have equivalent effect (R1=R2=R3, r1=r2=r3) [8], the dominant genes and the recessive genes in garden pea are replaced by R and r, the results obtained are shown in Table 2.
The actual numbers of the F2 plants are 0, 8, 15, 5 and 50, which fitted with their expected value in 5 genotype categories in Table 2 [11]. This indicates that the old reds of seed coat of the variety are indeed determined by 3 pairs of nuclear genes, effect of which is additive, and nuclear genes are segregated normally.
According to the number of R gene, in the F2 there are 7 types of genotypes that show the ratio 1:6:(12 + 3):(8 + 12):15:6:1 representing binomial distribution. They would correspond with the phenotypes in grain colors in the subsequent hybrid generation in wheat: 1) white, 2) very light red, 3) light red, 4) medium red, 5) dark red, 6) very dark red, and 7) old red.
The phenotypes of the F2 grain colors in wheat: 1) In Nilsson-Ehle’s hybridization experiment, “the grains (the F2) produced by the F1 plants were medium
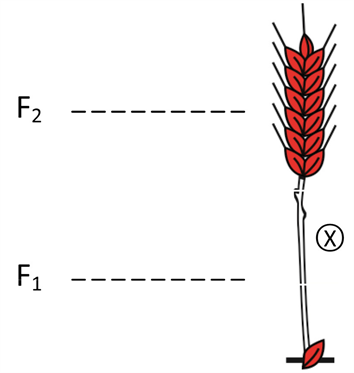
Figure 1. The F1 plants (r1R1r2R2r3R3).
Table 2. The F2 genotype categories in crossing with red grain wheat of 3 gene pairs.
red [6]”. 2) When a parent was red seed variety with a single gene pair, all the seeds produced by the F1 plants were lighter red1. The process may be diagrammed as follows (Figure 2).
So, the color of the grains (F2) produced by the F1 (rRrRrR) in Nilsson-Ehle’s hybridization experiment is uniformly medium red, including the grains with a genotype of rrrrrr, and does not segregate.
Like the old red of the F1 grains, the medium red of the F2 grains is also determined by mother’s genotype [6]. This is due to the pericarp inheritance. Tang Kai-dong, Yang Xue-ming and Yao Jin-bao etc. also found this feature in different cultivars of wheat [12][13][14]. This is neither maternal inheritance nor maternal effect.
There were two misunderstandings in current research community on the inheritance of grain colors in wheat. Some of them considered the inheritance of grain colors in wheat was the same as that of seed colors in garden pea. They were determined by genotype of 2n tissue of embryo. Like garden peas, the segregation of the grain colors in wheat occurred in the second hybrid generation [8][9][15][16]. This is fabricated. The other argued that, the pathway was from plants to plants to make the wheat propagation like viviparous species, which skipped forming of the pericarp originating from 2n tissues of mother’s body. Thereby, the grains were moved forward by one generation, and the F3 grains with different colors were said to be the traits of the F2 [1]. This is also improper.
3.2. Phenotypes of the F3 Grain Colors in Wheat Representing Segregation
The grains produced by the F1 plants above were sown and 78 F2 plants grew up, which distributed at random. They produced the F3 grains which represented different colors. For convenience reasons, according to the genotype classification in Table 2, the F2 plants were divided into 5 categories (Figure 3). In Figure 3, there are the actual number of the F2 plants, genotypes and their theoretical members, and as described in the previous section, standing for different color of the F3 grains.
The phenotypes of the F3 grain colors: 1) “The grains produced by the F2 plants were various shades of red and white” [6]. Nilsson-Ehle did not obtain the F2 plant producing pure white grains, however, from 5 F2 plants segregating the

Figure 2. The pericarp inheritance in wheat.

Figure 3. The F2 plants.
F3 plants producing red grains and white grains in the ratio 63:1, we knew that there could be such a F2 plant. 2) When a parent was red seed variety with a single gene pair, the F2 plants producing white grains, lighter red grains and red grains were in ratio 1:2:1, in another word, the ratio of the grains of different colors was 1:2:1 in the third hybrid generation2 [1]. So the segregation in the grain colors in Nilsson-Ehle’s hybridization experiment deferred a generation and in the third hybrid generation there were various shades of red grains and white grains. According to data in Table 2, their theoretical ratio should be 1:6:15:20:15:6:1, ranging from white to old red.
This is the inheritance of nuclear genes and another type of delayed inheritance which is different from that of silkworm eggs.
According to the relevant information in Figure 3 and the relevant theoretical values of the F2 plant distribution in Table 2, what can be calculated out was 6, 22, 15, 6, 1 plants producing light red, medium red, dark red, very dark red and old red grains out of 50 F2 plants producing red grains. Numbers of the F2 actual plants containing the calculated components were shown in Table 3.
Here the actual numbers of the F2 plants may differ from the real numbers. However, according to the theoretical distribution of the F2 plants corresponding to genotype in Table 2, there should be 7 types of actual plants, and when the number of experimental plants increases, they will approach the expected values and have certain fixed probability.
3.3. The Above Analysis Being Verified by the Segregation of the F4 Grain Colors
Mendel verified the Law of Segregation and the Law of Independence by the help of self-fertilization method [8]. The above analysis can be also verified by the method.
The F3 grains produced by 78 F2 plants were sown in plant-to-row nursery and 78 rows of the F3 plants grew up and produced the F4 grains. Like the previous section, according to genotype categories, the F3 plants would be divided into 5 categories (Figure 4).
Phenotypes of the F4 grain colors: “The F4 grains produced by offspring of F2 plants segregated reds and whites in the ratio 3:1, 15:1 and 63:1” [6]. But, these are not complete.
Table 3. The F3 grain phenotype and the F2 plant distribution.

Figure 4. Plant-to-row nursery of the F3 plants.
From the analysis of plant-to-row nursery of F3 plants we can obtain the following data:

The self-fertilized results are not in contradiction with the data obtained by Nilsson-Ehle. It is just that the latter described the wheat propagation as viviparous process, and the F4 grains were moved to the F3 generation. This is due to the limitation of historical conditions. It is the fact that these F2 plants produced the results by the help of twice self-fertilizing along with the pericarp inheritance, which fitted with their expected value in 5 genotype categories in Table 2. It shows that the above analysis on the variation of grain colors, pericarp inheritance and the additive effect of 3 gene pairs in wheat is correct.
Similar test can be accomplished in the F3 generation in garden pea and other crops, and it should appear in the F4 generation for grain color as wheat.
4. Conclusion and Discussion
Seeds of wheat are coated with 2n tissues of mother’s body [4][5]. In studies on the inheritance of grain colors in wheat there were two misunderstandings, in which scientists did not take this important feature into account, so that they did not perform the right analysis. In the study, self-fertilization method along with the pericarp inheritance has been adopted to analyze the inheritance in Nilsson-Ehle’s hybridization experiment in wheat. Because the outer coat in wheat seeds is 2n tissue of mother’s body, the grain colors are determined by mother’s genotype [6]. The color of the F1 grains in this experiment is old red. The color of the F2 grains is uniformly medium red and do not segregate. The segregation of the grain colors occurs in the third hybrid generation, which belongs to the delayed inheritance. It is wrong to think that the grain color in wheat segregates in the second hybrid generation in genetics works in the world [8][9][15][16][17]. The 7 traits containing 6 types of different red grains and 1 type of white grains in the third hybrid generation are the consequence of the inheritance of the additive effect of 3 gene pairs. It is the same as the inheritance of the additive effect of two pairs of independent dominant genes in the inheritance of qualitative characters [7][8], but is different from the cumulative effect resulted from adding unequal effect of many genes at many loci in the inheritance of quantitative traits [11]. Quantitative trait is often not a single unit characters or several unit character, but is a set of many unit character so that it involves numerous genes at fairly many loci, for example, more than 50 loci in corn chlorophyll [18]and hundreds or thousands loci in animal and plant biomass. The old red colors in the seed coat of the wheat variety used by Nilsson-Ehle only involve 3 gene pairs at 3 loci on 3D, 3A and 3B chromosomes [8][19], as far as the binomial distribution B(2n, p) is concerned, 2n = 6, p = 0.5, random variables have only 7 values and they may have determinate probabilities respectively. Recent studies have discovered that there are other loci on other chromosomes, alleles at which determine the red color of wheat grains. However, these loci are very limited and only produce 2n + 1 grades of grain colors. Mathematically, random variables are not a continuous distribution, but a discrete distribution [20]. The inheritance of grain colors in wheat still belongs to the categories of inheritance of qualitative traits. In addition, the strictly limiting condition of the multiple-gene hypothesis based on this experiment [8]limits the participation of many genes and makes “multiple gene” being only nominal. And it does not involve environmental effect which plays a large part in determining the phenotypes [9][11]. So the hypothesis is difficult to solve the continuous variations in the inheritance of quantitative characters.
Conflicts of Interest
The authors declare no conflicts of interest regarding the publication of this paper.
Cite this paper
Zhang, T.Z., Jia, X.M. and Xu, Z. (2018) An Alternative Analysis on Nilsson-Ehle’s Hybridization Experiment in Wheat-Theory of Dual Multiple Factors and Three Normal Distributions on Quantitative Inheritance (Continuation). Applied Mathematics, 9, 1005-1014. https://doi.org/10.4236/am.2018.98068
References
- 1. Klug, W.S., Cummings, M.R., Spencer, C.A., et al. (2017) Essentials of Genetics. 7th Edition, Higher Education Press, Beijing, 446-464
- 2. Suzuki, D.T., Griffiths, A.J.F. and Lewontin, R.C. (1990) An Introduction to Genetic Analysis. 2nd Edition, Shanxi People,s Press, Xian, 1002-1056. (In Chinese)
- 3. Nilsson-Ehle, H. (1909) Kreuzungsuntersuchungen an hafer und Weizen. Lunds Universitets Arsskrift, 7, 280-291.
- 4. Wang, W.H., Guan, X.L., Wang, D., Wang, R.Y. and Tian, Y.L. (2015) Botany. China Forestry Press, Beijing, 172-199. (In Chinese)
- 5. Ma, W.L., Wang, Y.F. and Li, H.Q. (2015) Botany. Higher Education Press, Beijing, 78-111. (In Chinese)
- 6. Zhao, H.Z., Wang, J.L. and Wang, J.Z. (1987) Crop Breeding. Agriculture Press, Beijing, 261-272. (In Chinese)
- 7. Jiao, T., Zhong, S.L., Gao, Q., et al. (1984) Plant Breeding. Hunan Science and Technology Press, Changsha, 8-59. (In Chinese)
- 8. Zhu, J., Liu, Q.C., Zhang, T.Z., et al. (2014) Genetics. 3rd Edition, Agriculture Press, Beijing, 65-339. (In Chinese)
- 9. Crow, J.F. (1976) Genetics Notes. 7th Edition, Burgess Publishing Company, Minneapolis, 161-202.
- 10. Liu, C.D. (2017) Genetics. 2nd Edition, Higher Education Press, Beijing, 446-464. (In Chinese)
- 11. Zhang, T.Z., Jia, X.M. and Xu, Z. (2016) The Limitations of Polygenic Hypothesis and Theorizing about Dual Multiple Factors and Three Normal Distributions. Applied Mathematics, 7, 912-919. https://doi.org/10.4236/am.2016.79081
- 12. Tang, K.D., Zhu, S.W., Bai, J.J., et al. (2011) Preliminary Studies on Inheritance of Seed Color in Black-Seeded Wheat. Agricultural Science and Technology, 12, 1141-1143.
- 13. Yang, X.M., Zhou, M.P., Yao, J.B., et al. (2014) Inheritance of Grain Coat Color in Black-Grained Wheat Ning 0726. Jiangsu Journal of Agricultural Science, 30, 721-723.
- 14. Yao, J.B., Yao, G.C., Yang, X.M., et al. (2007) Inheritance of Seed Coat Color in Two Wheat Cultivars. Jiangsu Journal of Agricultural Science, 23, 721-723 + 267-269.
- 15. Dai, Z.H., Ya, F., Ding, Y., et al. (2016) Genetics. Third Edition, Higher Education Press, Beijing, 126-147. (In Chinese)
- 16. Lu, L.D. (2017) Geneal Genetics. Science Press, Beijing, 109-129. (In Chinese)
- 17. Ayala, F.J. and Kiger, J.A. (1988) Modern Genetics. Hunan Science and Technology Press, 600-629. (In Chinese)
- 18. Fang, Z.X. (1984) General Genetics. Science Press, Beijing, 64-351. (In Chinese)
- 19. Zhu, Z.W., Li, J.H., Tong, H.W., et al. (2016) Progress on Inheritance of Red Grain Trit of Wheat. Hubei Agricultural Science, 55, 6321-6323.
- 20. Shen, Z., Xie, S.Q. and Pan, C.Y. (2011) Probability Theory and Mathematical Statistics. 4th Edition, Higher Education Press, Beijing, 20-120. (In Chinese)
NOTES
1Northwest A & F University, Li Z. D., Genetics lecture note for postgraduates (1978).
2Northwest A & F University, Li Z. D., Genetics lecture note for postgraduates (1978).
上一篇:Ordered Rate Constitutive Theo 下一篇:Modeling and Numerical Solutio
最新文章NEWS
- Ordered Rate Constitutive Theories for Non-Classical Thermoviscoelastic Fluids with Internal Rotatio
- Modeling and Numerical Solution of a Cancer Therapy Optimal Control Problem
- Extended Wiener Measure by Nonstandard Analysis for Financial Time Series
- Methodology for Constructing a Short-Term Event Risk Score in Heart Failure Patients
- Application of Conjugate Gradient Approach for Nonlinear Optimal Control Problem with Model-Reality
- On the Dynamics of Transition of a Classical System to Equilibrium State
- On the ECI and CEI of Boron-Nitrogen Fullerenes
- Binomial Hadamard Series and Inequalities over the Spectra of a Strongly Regular Graph
推荐期刊Tui Jian
- Chinese Journal of Integrative Medicine
- Journal of Genetics and Genomics
- Journal of Bionic Engineering
- Pedosphere
- Chinese Journal of Structural Chemistry
- Nuclear Science and Techniques
- 《传媒》
- 《中学生报》教研周刊
热点文章HOT
- An Alternative Analysis on Nilsson-Ehle’s Hybridization Experiment in Wheat —Theory of Dual Multiple
- On Existence of Periodic Solutions of Certain Second Order Nonlinear Ordinary Differential Equations
- Ordered Rate Constitutive Theories for Non-Classical Thermoviscoelastic Fluids with Internal Rotatio
- Application of Conjugate Gradient Approach for Nonlinear Optimal Control Problem with Model-Reality
- Fuzzy Modelling for Predicting the Risk of Recurrence and Progression of Superficial Bladder Tumors
- Integer Wavelet-Based Image Interpolation in Lifting Structure for Image Resolution Enhancement
- The Invulnerability of Directed Interdependent Networks with Multiple Dependency Relations
- Extended Wiener Measure by Nonstandard Analysis for Financial Time Series